Омега-3 полиненасыщенные жирные кислоты в ревматологии: І. Теоретические основыКлючевые слова: полиненасыщенные жирные кислоты, эйкозаноиды, цитокины, ревматические болезни. Резюме. Омега-3 полиненасыщенные жирные кислоты (ПНЖК), а именно эйкозапентаеновая (ЭПК) и докозагексаеновая кислоты, содержатся в жире морских рыб и некоторых морепродуктах. Поступая с пищей, они соперничают с омега-6 ПНЖК, прежде всего арахидоновой кислотой (АК), за место в мембранных фосфолипидах. В последующем ЭПК вступает в конкурентные отношения с АК как субстрат для циклооксигеназного и липоксигеназного путей метаболизма. Получаемые из ЭПК простагландины и лейкотриены имеют значительно более низкую биологическую активность по сравнению с метаболитами АК. Складывающийся в результате баланс эйкозаноидов имеет противовоспалительную и антитромботическую направленность. Путем угнетения синтеза из АК лейкотриенов 4-ой серии омега-3 ПНЖК снижают также выработку мононуклеарными клетками провоспалительных цитокинов – фактора некроза опухоли, интерлейкинов 1, 2 и 6. Эти данные подтверждены в исследованиях на животных и, особенно убедительно, на здоровых людях, которые получали диету, обогащенную омега-3 ПНЖК. Положительное влияние пищевых омега-3 ПНЖК на профиль синтезируемых эйкозаноидов и цитокинов, а также на денатурантную стабильность эндогенных белков может быть основанием для их применения в лечении иммунозависимых хронических воспалительных заболеваний. Ревматоидный артрит (РА), другие хронические артропатии, диффузные болезни соединительной ткани являются практически неизлечимыми заболеваниями, при которых больные вынуждены длительно, зачастую непрерывно принимать противовоспалительные препараты. Долгое время вполне понятный и широко распространенный интерес пациентов к фитотерапии, разного рода диетам и пищевым добавкам, "мегадозной" витаминотерапии как альтернативе или хотя бы дополнению к традиционным фармакологическим препаратам наталкивался на обоснованный скептицизм в среде практикующих врачей и научном сообществе. Между тем в последние два десятилетия в контролируемых исследованиях получены данные, свидетельствующие о реальном патогенетическом воздействии и клинической эффективности по крайней мере некоторых методов нетрадиционной фармакотерапии. Одним из наиболее теоретически обоснованных направлений, развитие которого завершилось промышленным выпуском препаратов, является применение в лечении хронических воспалительных ревматических заболеваний омега-3 полиненасыщенных жирных кислот (ПНЖК). История вопроса. Толчком к проведению научных работ в этой области послужили эпидемиологические исследования, продемонстрировавшие необычайно низкую распространенность инфаркта миокарда и атеросклероза у эскимосов острова Гренландия по сравнению с сопоставимыми по полу и возрасту группами населения континентальной Европы (Dyerberg J. et al., 1975; Kromann N., Green A., 1980; Kromhout D. et al., 1985). Bang H.O. et al. в 1975 г. предположили, что это может быть связано с высоким потреблением с пищей рыбьего жира, содержащего омега-3 жирные кислоты. Повышенный интерес к взаимосвязи между количеством потребляемых морепродуктов и сердечно-сосудистыми заболеваниями, подтвердившейся и на жителях Японии (Hirai A. et al., 1980; Kagawa Y. et al., 1982), на время затмил установленный в то же время (Kromann N., Green A., 1980) факт, что у эскимосов очень редко встречаются аутоиммунные и воспалительные заболевания типа бронхиальной астмы, РА, псориаза, сахарного диабета І типа и полностью отсутствует рассеянный склероз. Объяснения этого также строились на гипотезе о потенциально положительном эффекте омега-3 ПНЖК (Horrobin D.F., 1987). Действительно, у эскимосов наблюдался более высокий, чем в других популяциях, сывороточный уровень эйкозапентаеновой кислоты (ЭПК), а также дигомо-g-линоленовой (эйкозатриеновой) кислоты, и при этом - более низкие уровни арахидоновой (эйкозатетраеновой) (АК) и линолевой кислот из семейства омега-6. Хотя и оказалось, что различия в соотношении уровня этих кислот являются следствием не только диеты, но и генетически обусловлены в этой популяции (Horrobin D.F., 1987), но дальнейшие наблюдения представили убедительные аргументы в пользу того, что метаболиты омега-3 кислот являются для организма более полезными и способны влиять на течение воспалительных и имунных реакций. Номенклатура ПНЖК. Из более 200 существующих в природе жирных кислот в тканях человека и животных обнаруживаются около 70, а практически значимое распространение имеют немногим более 20 (Климов А.Н., Никульчева Н.Г., 1995). Примерно 3/4 жирных кислот являются ненасыщенными, то есть содержат двойные связи. В соответствии с сокращенной систематической номенклатурой они обозначаются двумя цифрами, например, линолевая С 18:2, где первая цифра указывает число углеродных атомов в молекуле, а вторая – число ненасыщенных связей. По новой номенклатуре указывается очередность атома углерода с первой двойной связью по отношению к углероду концевой метильной группы (омега-положение; от w - последней буквы греческого алфавита, символа конца) и рациональное название кислоты греческого происхождения, первая часть которого обозначает число атомов углерода (октадека – 18, эйкоза – 20, докоза – 22 и т.д.), а вторая – количество двойных связей – "ен" (ди – 2, три – 3, тетра – 4, пента – 5, гекса – 6). Характеристика некоторых наиболее физиологически важных ненасыщенных жирных кислот приведена в таблице 1. Во многих зарубежных публикациях концевой (метильной группы) углерод жирной кислоты обозначают не греческой буквой w, а латинской n. Таблица 1. Номенклатура и характеристика некоторых физиологически важных ненасыщенных жирных кислот.
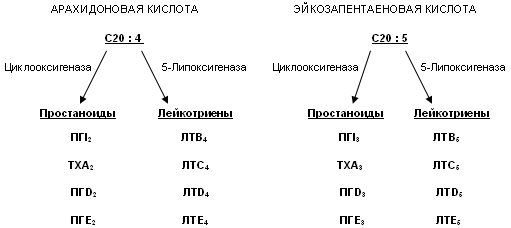
Сравнительная физиология обмена омега-3 и омега-6 ПНЖК. Клетки млекопитающих способны синтезировать только омега-9 жирные кислоты, которые содержат не более трех двойных связей и не далее чем у 9-го атома углерода от метильного конца. Незаменимые (эссенциальные) омега-6 и омега-3 ПНЖК поступают в организм только с пищей, поскольку в клетках млекопитающих нет ферментов, катализирующих введение двойных связей в цепь жирных кислот далее 9-го атома углерода. При этом омега-6 ПНЖК – линолевая, g-линоленовая и арахидоновая содержатся в растительных маслах и в фосфолипидах животных, а омега-3 ПНЖК – эйкозапентаеновая и докозагексаеновая (ДГК) – в фитопланктоне и жире поедающих его морских рыб (лосось, макрель, сардина, сельдь и др.). Еще одна кислота семейства омега-3 - a-линоленовая обнаруживается в грецких орехах, льняном семени, соевом масле (DeLuca P. et al., 1995; James M.J. et al., 2000). Поступающие с пищей ПНЖК уже в энтероцитах ресинтезируются в фосфолипиды, которые структурируются в липопротеиды высокой плотности и переносятся непосредственно к клеткам всех тканей. В момент ассоциации липопротеидов с клетками более полиеновая экзогенная жирная кислота переходит из фосфолипида липопротеида в фосфолипид клеточной мембраны, а менее полиеновая эндогенная кислота — в обратном направлении. Таким пассивным (нерегулируемым) путем клетки поглощают омега-6 ПНЖК. Омега-3 ПНЖК поступают в клетки более сложным, рецепторным путем: после этерификации холестерином они в составе липопротеидов низкой плотности поглощаются посредством эндоцитоза через апоВ-100 рецепторы (Титов В.Н., 2001). В основном поглощают омега-3 ПНЖК два вида клеток – 1) высокодифференцированные и 2) специализированные клетки рыхлой соединительной ткани. Первые используют эссенциальные ПНЖК как предшественники и синтезируют из них более сложные кислоты, которые, встраиваясь в мембрану, во многом определяют специфичную функцию клеток (клетки сетчатки, гломерулярной мембраны почек, нейроциты, сперматозоиды и др.). Вторые (эндотелий, гладкомышечные клетки, фибробласты, тромбоциты, нейтрофилы, моноциты и макрофаги) синтезируют из омега-6 (главным образом, АК) и омега-3 ПНЖК (в основном, ЭПК) биологически активные вещества — простагландины, тромбоксаны и лейкотриены (Титов В.Н., 2001; Linn D.S. et al., 1998). АК является предшественницей так называемой 2-ой серии простагландинов и 4-ой серии лейкотриенов, а ЭПК – соответственно 3-ей и 5-ой серий (см. рис.1). Рис. 1. Эйкозаноиды, получаемые из арахидоновой (семейство w-6) и эйкозапентаеновой (семейство w-3) кислот.
Эйкозаноидный механизм противовоспалительного действия омега-3 ПНЖК. Функциональные свойства эйкозаноидов, получаемых из АК и ЭПК, являются во многом противоположными. Так, ПГE2 повышает функциональную активность полиморфноядерных нейтрофилов, а ПГE3 снижает ее. Если TXA2 индуцирует выраженное сужение сосудов, активацию нейтрофилов и тромбоцитов, то TXA3 - слабый вазоконстриктор, ингибирует агрегацию тромбоцитов, уменьшает биологические эффекты этих клеток (Belch J.J. et al., 1988; Raederstorff D. et al., 1996). Напротив, ПГI3 и простациклин (ПГI2) сопоставимы по своей антикоагулянтной активности и вазодилататорной потенции (Fischer S., Weber P., 1984). Полагают, что именно это различие соотношений биологической активности тромбоксанов и простациклинов 2-ой и 3-ей серии ответственно за удлинение времени кровотечения и ослабление агрегации тромбоцитов, наблюдающееся у эскимосов Гренландии (Dyerberg J., Bang H.O., 1979). Получаемые из АК лейкотриены обладают очень мощными провоспалительными свойствами (Dyerberg J., Bang H.O., 1979; James M.J. et al., 2000). ЛТB4, продуцируемый нейтрофилами, является активным хемоаттрактантом, он усиливает миграцию в очаг воспаления и адгезию нейтрофилов, моноцитов и макрофагов, вызывает дегрануляцию нейтрофилов, повышает сосудистую проницаемость. Другие лейкотриены - C4, D4 и E4 (выделяются главным образом эозинофилами), кроме увеличения сосудистой проницаемости, индуцируют спазм гладких мышц в сосудистой стенке и бронхах. Основной продукт липоксигеназного пути метаболизма ЭПК – ЛТB5 имеет лишь от 1% до 10% разных видов биологической активности ЛТB4 (табл. 2) и, кроме того, подавляет синтез ЛТB4. В то же время продуцируемые эозинофилами ЛТC5, ЛТD5 и ЛТE5 по способности вызывать спазм гладкой мускулатуры близки лейкотриенам 4-ой серии, хотя и подавляют продукцию последних (Lee T.H., Austen K.F., 1986). С учетом этих данных в научных исследованиях для оценки биохимических последствий диеты, обогащенной омега-3 ПНЖК, чаще всего определяют уровень в крови именно ЛТB4 и ЛТB5. Таблица 2. Сравнительная биологическая активность ЛТB4 и ЛТB5.
Примечание. Все виды активности ЛТB4 условно приняты за 100. Таким образом, в целом эйкозаноиды – производные омега-6 ПНЖК обладают провоспалительными, вазоконстрикторными и проагрегантными свойствами, обеспечивая филогенетически закрепленные защитные реакции – воспаление и остановку кровотечения. Эйкозаноиды, синтезируемые из омега-3 ПНЖК, имеют противовоспалительное и антитромботическое действие как противовес биологическим эффектам метаболитов АК. Если содержание в пище омега-3 ПНЖК увеличивается, ЭПК и ДГК соперничают с АК за место в мембранных фосфолипидах, частично вытесняют последнюю и в дальнейшем вступают в конкурентные отношения с ней в качестве субстрата для дальнейших биохимических превращений, в частности в циклооксигеназном и липоксигеназном путях. В результате равновесие смещается в сторону синтеза эйкозаноидов с противовоспалительными и дезагрегантными свойствами. Уменьшение содержания в клеточных мембранах АК и сопутствующее снижение синтеза соответствующих эйкозаноидов под влиянием диеты, обогащенной омега-3 ПНЖК (главным образом в виде рыбьего жира или его концентратов), продемонстрировано в многочисленных исследованиях (Enders S. et al., 1989; Brouard C., Pascaud M., 1990; Hardardottir I., Kinsella J.E., 1992; Fritsche K.L. et al., 1993; Black J.M., Kinsella J.E., 1993; Yaqoob P., Calder P.C., 1995). При этом доля ЭПК и ДГК в мембранных жирных кислотах возрастала, а производство простагландинов и тромбоксанов 3-ей серии и лейкотриенов 5-ой серии увеличивалось (Brouard C., Pascaud M., 1990; Chapkin R.S. et al., 1990; Grimminger F. et al., 1993). Обогащение пищевого рациона омега-3 ПНЖК приводило также к подавлению генерации нейтрофилами и моноцитами еще одного воспалительного медиатора-хемоаттрактанта - фактора активации тромбоцитов (Lee T.H. et al., 1985; Sperling R.L. et al., 1987; Schmidt E.B. et al., 1991; Sperling R.I., 1991). Интересно, что и семейство омега-6 кислот включает в себя одну ПНЖК – дигомо-g-линоленовую, метаболиты которой имеют ярко выраженную противовоспалительную направленность, подобную таковой производных семейства омега-3 ПНЖК. В условиях внутриклеточного дефицита омега-3 ПНЖК с пятью и шестью двойными связями и омега-6 ПНЖК с четырьмя связями высокодифференцированные клетки и специализированные клетки рыхлой соединительной ткани компенсаторно начинают сами синтезировать из эндогенной омега-9 олеиновой жирной кислоты дигомо-g-линоленовую ПНЖК (омега-6), которая имеет только три двойные связи (Климов А.Н., Никульчева Н.Г., 1999). Естественно, что физико-химические свойства триеновой ПНЖК существенно отличаются от эссенциальных АК, ЭПК и ДГК. Использование дигомо-g-линоленовой кислоты высокодифференцированными клетками при сборке фосфолипидов имеет в целом нежелательные последствия: увеличивается микровязкость мембраны, снижается отрицательный заряд на ее поверхности, что может приводить к дисфункции мембранных белков и самих клеток (Титов В.Н., 2001). В то же время в специализированных соединительнотканных клетках образуются метаболиты дигомо-g-линоленовой кислоты, которым приписывается благоприятное противовоспалительное действие. Эта кислота конкурирует с АК за окислительные ферменты, таким образом сокращая производство циклооксигеназных продуктов АК (Kernoff P.B.A. et al., 1977). В циклооксигеназном пути метаболизма из дигомо-g-линоленовой кислоты образуется ПГE1 (Manku M.S. et al., 1986). Вместе с другими простагландинами ПГE1 стимулирует развитие кардинальных признаков воспаления, однако действие его на клетки воспаления главным образом подавляющее. ПГE1 увеличивает внутриклеточное содержание цАМФ и это приводит к ослаблению хемотаксиса, прилипания полиморфноядерных лейкоцитов к сосудистой стенке и высвобождения ими лизосомальных ферментов. Подобное депрессивное влияние оказывает ПГE1 и на лимфоциты (Rogers T.H., 1985; Belch J.J.F., Muir A., 1998). Полагают, что ПГE1 выполняет функцию отрицательной обратной связи при хроническом воспалительном процессе, первоначально помогая развитию кардинальных признаков воспаления, а затем оказывая супрессивное действие (Belch J.J.F., Muir A., 1998). Сама дигомо-g-линоленовая кислота не может быть преобразована в воспалительные лейкотриены (Hammarstrom S., 1981) и, кроме того, формирует производные, которые блокируют метаболические превращения АК в лейкотриены 4-ой серии (Voorlees, 1993). Следовательно, увеличение поступления в организм этой ПНЖК или g-линоленовой кислоты (которая способна в клетках метаболизироваться в дигомо-g-линоленовую) может конкурентно ингибировать выработку 2-ой серии простагландинов и 4-ой серии лейкотриенов и таким образом подавлять активность хронических воспалительных заболеваний типа РА (Jantti J. et al., 1989; Oxholm P. et al., 1992). Обнаружено также снижение агрегации лейкоцитов под влиянием добавления в пищу g-линоленовой кислоты (Maple С. et al., 1998). Приведенные данные явились основанием для многих центров, занимающихся фитотерапией, рекомендовать включать в пищевые добавки некоторые растения (примула аптечная, огуречник лекарственный и др.) или их масла, содержащие дигомо-g-линоленовую кислоту. Цитокиновый механизм противовоспалительного действия омега-3 ПНЖК. Биологические эффекты омега-3 ПНЖК не исчерпываются только описанными механизмами. Увеличение поступления этих кислот в организм сопровождается также снижением синтеза мононуклеарными клетками провоспалительных и иммунорегуляторных цитокинов (Enders S. et al., 1989; Schmidt E.B. et al., 1991). Избыточная продукция цитокинов, прежде всего фактора некроза опухоли a (ФНО-a), интерлейкина 1 (ИЛ-1) и интерлейкина 6 (ИЛ-6), ассоциируется со многими воспалительными ревматическими заболеваниями (Насонов Е.Л., Самсонов М.Ю., 1997; Al-Janadi M. et al., 1993). Цитокины представляют собой растворимые белковые медиаторы, продуцируемые многими мононукларными клетками вследствие их активации специфическими стимулами. Цитокины влияют на деятельность эффекторных клеток, связываясь со специальными рецепторами, экспрессированными на их поверхности. Первым в каскаде цитокинов медиатором, выделяемым активированными моноцитами и макрофагами, является ФНО-a. Он может также продуцироваться лимфоцитами, натуральными киллерами, нейтрофилами и тучными клетками. ФНО-a стимулирует нейтрофилы, моноциты и макрофаги, повышает экспрессию молекул адгезии на поверхности нейтрофилов и эндотелиоцитов и этим способствует увеличению проницаемости эндотелиальных белков, активирует функцию Т- и В-клеток, усиливает контроль за антигенами комплекса гистосовместимости и инициирует синтез других провоспалительных цитокинов – ИЛ-1 и ИЛ-6. ИЛ-1 – второй цитокин, освобождаемый в ответ на воспалительные стимулы, включая ФНО-a. Кроме моноцитов и макрофагов, он может быть произведен также другими разновидностями клеток типа эндотелиоцитов, кератиноцитов, В-клеток. ИЛ-1 дублирует многие провоспалительные клинико-метаболические эффекты ФНО-a, включая лихорадку, сонливость, недомогание, снижение аппетита, потерю в весе, снижение синтеза альбумина и повышение синтеза гепатоцитами белков острой фазы (С-реактивного протеина, орозомукоида, трансферрина, протромбина, фибриногена и других), нейтрофилез и лимфопению, стимуляцию продукции других цитокинов, в том числе и ФНО-a (табл. 3). Кроме того, он потенциирует активность прокоагулянтов и повышает выработку ингибитора активатора плазминогена и эндотелина, а также эйкозаноидов. В ответ на действие ИЛ-1 и ФНО-a мононуклеарами (среди прочих – и Т-клетками) вырабатывается ИЛ-6, играющий важную роль в воспалении, но имеющий более узкий спектр действия по сравнению с первыми двумя ключевыми цитокинами (Насонов Е.Л., Самсонов М.Ю., 1997; Белобородов В.Б., 2001; Endres S. et al., 1995; Feldman M. et al., 1996). Следовательно, выделение цитокинов в межклеточное пространство и в кровоток обеспечивает сигнальные функции, связанные с формированием генерализованной воспалительной реакции. Таблица 3. Биологические эффекты провоспалительных цитокинов.
В исследованиях на животных получено много доказательств, что скармливание растительных масел и рыбьего жира, богатых омега-3 ПНЖК, изменяет, чаще в сторону снижения, производство ФНО-a, ИЛ-1, ИЛ-6 и ИЛ-2, хотя имеется много противоречащих наблюдений (Endres S. et al., 1995; Calder P.C., 1997). Наиболее вероятно, несоответствие результатов вытекает из различий использовавшихся протоколов экспериментов (количество жира и длительность приема, характер сравнительной диеты, разновидность используемого животного, способы выделения макрофагов, стимуляторы синтеза цитокинов и их концентрация, способы культивирования клеток и т. п.) и типов исследовавшихся клеток - продуцентов цитокинов (альвеолярные, перитонеальные макрофаги, купферовские клетки, мононуклеары периферической крови). Исследования на людях дали более последовательные результаты: в нескольких работах продемонстрировано, что обогащение диеты здоровых добровольцев омега-3 ПНЖК приводит к уменьшению выработки ИЛ-1, ИЛ-6, ФНО-a и ИЛ-2 мононуклеарными клетками периферической крови (Enders S. et al., 1989; Meydani S.N. et al., 1991; Molvig J. et al., 1991; Virella G. et al., 1991; Calder P.C., Newsholme E.A., 1992; Cooper A.L. et al., 1993; Meydani S.N. et al., 1993; Caughey G.E. et al., 1996). Endres S. et al. (1989) первыми показали, что у здоровых людей употребление с пищей повышенных количеств омега-3 ПНЖК уже через 6 недель приводит к достоверному снижению продукции мононуклеарами ИЛ-1a и ИЛ-1b, причем выработка цитокинов оставалась угнетенной и спустя 10 недель после возврата к обычной диете. Это объясняют длительным периодом "вымывания", способностью ЭПК и ДПК пролонгированно инкорпорироваться в мембрану циркулирующих клеток. Лишь через 20 недель после окончания приема высоких доз омега-3 ПНЖК продукция ИЛ-1 возвращалась к исходному уровню. Подобным образом, но менее выраженно, изменялась и продукция ФНО-a. В более поздней работе этих же авторов (Endres S. et al., 1993) доказано супрессивное влияние аналогичной диеты и на выработку ИЛ-2. Эти и упомянутые выше другие исследователи приводят цифры снижения уровня ИЛ-1 в основном в пределах 30-85%, ФНО-a - до 40%. Caughey G.E. et al. (1996) показали наличие корреляции между содержанием ЭПК в мононуклеарных клетках и продукцией ИЛ-1 и ФНО-a. Подавление синтеза цитокинов омега-3 жирными кислотами, вероятно, достигается на уровне транскрипции генетической информации, на что указывают сниженные уровни информационной (матричной) РНК (Endres S. et al., 1995). В последние годы большее значение придается эйкозаноид-опосредованному механизму (Calder P.C., 1997). Выработка цитокинов и продукция эйкозаноидов находятся в тесной функциональной взаимосвязи. С одной стороны, как видно из таблицы 3, цитокины активируют фосфолипазу А2 и синтез эйкозаноидов, с другой стороны, циклооксигеназные и липоксигеназные метаболиты арахидоновой кислоты способны влиять на выработку провоспалительных цитокинов. Так, высокие концентрации ПГE2 ингибируют выработку ФНО-a моноцитами и макрофагами путем увеличения внутриклеточного уровня циклического АМФ (Renz H. et al., 1988; Endres S. et al., 1991). Напротив, низкие концентрации ПГE2 стимулируют выработку циклического ГМФ и в результате увеличивают продукцию ФНО-a (Kovacs E.J. et al., 1988). Имеются сообщения о том, что ПГE2 угнетает производство ИЛ-1, хотя эти данные не столь однозначны. ПГE1 и ПГE2 подавляют выработку ИЛ-2 T-лимфоцитами (Paliogianni F. et al., 1993). Липоксигеназный продукт ЛТB4 увеличивает производство ФНО-a моноцитами (Gagnon L. et al., 1989), а при лечении ингибиторами липоксигеназы концентрация ФНО-a снижается (Schade U.F. et al., 1989). Как и в отношении ПГE2, сведения о влиянии ЛТB4 и ЛТC4 на выработку ИЛ-1 противоречивы. В то же время известно, что ЛТB4 увеличивает продукцию ИЛ-6 и интерферона-g человеческими моноцитами и лимфоцитами (Rola-Pleszczynski M. et al., 1983; Poubelle P. et al., 1991). Таким образом, в целом ПГE2 подавляет производство ФНО-a, ИЛ-1, ИЛ-6 и ИЛ-2, в то время как лейкотриены 4-ой серии увеличивают выработку этих цитокинов. При частичном вытеснении АК из фосфолипидов мембран кислотами семейства омега-3 объем синтезируемых из АК эйкозаноидов ПГE2 и, особенно, ЛТB4 cнижается, вследствие чего уменьшается и выработка провоспалительных цитокинов. По-видимому, это – главный механизм, лежащий в основе ингибирующего влияния омега-3 ПНЖК на продукцию цитокинов с провоспалительными свойствами. Новую, дополнительную гипотезу для объяснения механизма возможного положительного влияния омега-3 ПНЖК на течение хронических аутоиммунных воспалительных болезней предложили Saso L. et al. (1999). Авторы исходили из того, что при этих заболеваниях имеет место денатурация и гликозилирование эндогенных сывороточных белков (как острофазовых, так и неострофазовых), что является одной из причин их антигенности (Silvestrini B. et al., 1989; Saso L. et al., 1993а, 1993б), поддерживающей и усиливающей аутоиммунный процесс. Сравнительное исследование различных солей, кислот и препаратов показало, что in vitro антиденатурантная активность ПНЖК рыбьего жира в отношении человеческого альбумина выше, чем таковая известных антиденатурантных лекарственных средств (бендазака и биндарита) и нестероидных противовоспалительных препаратов (фенилбутазона и индометацина). В эксперименте на животных омега-3 ПНЖК, применяемые орально, защищали сывороточные белки от температурной денатурации больше, чем бендазак (Saso L. et al., 1999). Следовательно, стабилизация эндогенных белков – еще один потенциально выгодный результат применения омега-3 ПНЖК при ревматических заболеваниях. Таким образом, омега-3 ПНЖК, выступая в качестве субстрата для окисления в циклооксигеназном и липоксигеназном путях, способствуют выработке простагландинов и лейкотриенов с противовоспалительными и антитромботическими свойствами. Увеличение содержания этих кислот в фосфолипидах клеточных мембран сопровождается угнетением продукции цитокинов, играющих ключевую роль в развитии хронических воспалительных процессов. Итоговый метаболический результат в виде снижения потенциала воспаления и дезагрегантно-гипокоагуляционных сдвигов, а также, вероятно, антиденатурантный эффект дают основания рассчитывать на положительный клинический эффект применения омега-3 ПНЖК при ревматических заболеваниях. Исследования на животных и здоровых людях свидетельствуют, что реальным путем увеличения содержания в организме и, в частности, в клеточных мембранах омега-3 ПНЖК является пероральное употребление определенных количеств рыбьего жира или соответствующих препаратов. ЛИТЕРАТУРА:
Яременко О.Б. Резюме. Омега-3 поліненасичені жирні кислоти (ПНЖК), а саме ейкозапентаәнова (ЕПК) і докозагексаәнова кислоти, містяться в жиру морських риб і деяких морепродуктах. Надходячи з ҝжею, вони конкурують з омега-6 ПНЖК, насамперед арахідоновою кислотою (АК), за місце в мембранних фосфоліпідах. Надалі ЕПК вступаә в конкурентні відношення з АК як субстрат для циклооксигеназного і ліпоксигеназного шляхів метаболізму. Простагландини і лейкотріәни, отримані з ЕПК, мають значно нижчу біологічну активність у порівнянні з метаболітами АК. Баланс ейкозаноҝдів, що утвоюәться в результаті, маә протизапальну й антитромботичну спрямованість. Шляхом пригнічення синтезу з АК лейкотріәнів 4-оҝ серіҝ омега-3 ПНЖК знижують також продукцію мононуклеарними клітинами прозапальних цитокінів - фактора некроза пухлини, інтерлейкінів 1, 2 і 6. Ці дані підтверджені в дослідженнях на тваринах і, особливо переконливо, на здорових людях, що одержували діәту, збагачену омега-3 ПНЖК. Позитивний вплив харчових омега-3 ПНЖК на профіль синтезованих ейкозаноҝдів і цитокінів, а також на денатурантну стабільність ендогенних білків може бути підставою для ҝхнього застосування в лікуванні імунозалежних хронічних запальних захворювань. Ключові слова: поліненасичені жирні кислоти, ейкозаноҝди, цитокіни, ревматичні хвороби.
O.B. Yaremenko Summary. Omega-3 polyunsaturated fatty acids (PUFAs), namely eicosapentaenoic (EPA) and docosahexaenoic acid, are contained in fat of sea fishes and some sea's products. Acting with food, they compete with omega-6 PUFAs, first of all by arachidonic acid (АA), for a place in membrane phospholipids. In subsequent EPA enters the competitive relations with AA as substratum for metabolism by the cyclooxygenase and lipoxygenase pathways. Received from EPA prostaglandins and leukotrienes have much lower biological activity in comparison with AA-derived metabolites. The eicosanoids balance, developing in result, has anti-inflammatory and anti-thrombotic orientation. Inhibiting synthesis of 4-series leukotrienes from АA, omega-3 PUFAs reduce also production by mononuclear cells of pro-inflammatory cytokines - tumour necrosis factor, interleukin 1, 2 and 6. These data are confirmed in animal studies and, especially convincingly, in studies on healthy people which received a omega-3 PUFAs enriched diet. The positive influence of dietary omega-3 PUFAs on a profile synthesized eicosanoids and cytokines and also on denaturant stability of endogenous proteins can be the basis for their application in treatment of immuno-dependent chronic inflammatory diseases. Key words: polyunsaturated fatty acids, eicosanoids, cytokines, rheumatic diseases. comments powered by Disqus |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||